Nuttige opmerkingen over het mechanisme van overschrijding
Handige notities over het mechanisme van oversteken!
Volgens Morgan wordt de koppeling veroorzaakt doordat gekoppelde genen worden uitgevoerd door hetzelfde chromosoom. Hij wees er ook op dat het fenomeen van volledige koppeling zelden voorkomt omdat soms de gekoppelde genen de neiging vertonen om te scheiden tijdens meiose. Op deze manier kan de kruising worden gedefinieerd als "een mechanisme van de recombinatie van de genen als gevolg van de uitwisseling van chromosomale segmenten op het moment van koppelen."

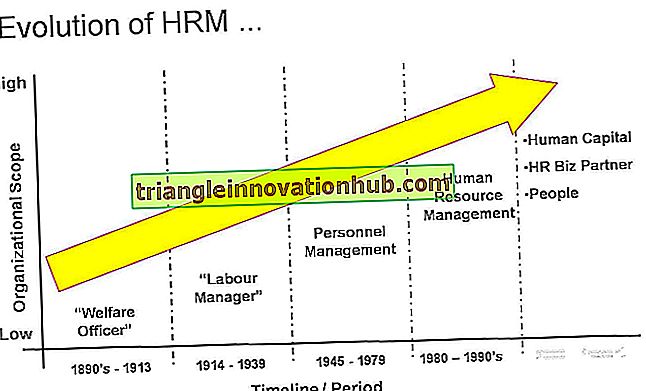
Afbeelding met dank aan: wikilectures.eu/images/1/1e/Crossing-over-2.jpg
De cross-over tussen gekoppelde genen maakt hun recombinatie mogelijk tijdens meiose. Het mechanisme van cross-over is als volgt:
Mechanisme van oversteken:
De figuur laat zien dat kruising, leidend tot recombinatie van gekoppelde genen, het gevolg is van uitwisseling van secties van homologe chromosomen. Bij meiose komen de homologe (maternale en vaderlijke) chromosomen bij elkaar en paren of synaps tijdens profase. De koppeling is opmerkelijk precies en wordt duidelijk veroorzaakt door wederzijdse aantrekking van de delen van de chromosomen die vergelijkbaar of homoloog zijn omdat ze allellische genen bevatten.
Verder, tijdens de overgang tussen de pachytene en de diplotene van de profase van meiose, verdelen de gepaarde chromosomen elk in twee chromatiden, zodat de bivalent nu is samengesteld uit vier chromatiden. Ongeveer op het moment dat de chromosomen voor het eerst gezien werden verdeeld, stellen de chromatiden één of meer uitwisselingen vast, of Chiasmata, per bivalent.
Bij elk chiasma zijn twee van de chromatiden gebroken en vervolgens opnieuw samengevoegd, zodat de nieuwe chromatiden nu worden gevormd uit secties van de oorspronkelijke. Door chiasma-vorming worden maternale en vaderlijke chromosomen niet overgedragen als ondeelbare eenheden; ze wisselen secties uit, en de nieuwe chromosomen die ontstaan als gevolg van meiose dragen genen die vóór meiose zich bevonden in verschillende leden van het paar homologe chromosomen. Janssens (1909) ontdekte voor het eerst het proces van chiasma-vorming. Dit is een zeer nauwkeurig proces, aangezien de twee chromatiden bij een chiasma exact equivalente segmenten uitwisselen, zodat, met zeer zeldzame uitzondering, geen van de chromiden winsten verliest of genen verliest.

Afwezigheid van oversteken bij Drosophila-mannetjes:
In de meeste organismen vindt recombinatie van gekoppelde genen zowel bij vrouwen als bij mannetjes plaats. Evenzo vindt in de meeste organismen de vorming van Chiasmata plaats in zowel vrouwelijke als mannelijke meiose. Maar in Drosophila is de situatie heel anders. In Drosophila vindt een kruising zelden of nooit plaats bij de man. In de zijderupsmot lijkt het echter afwezig te zijn bij de vrouwtjes.
Recombinatie en overschrijding:
De cytologische waarnemingen hebben één belangrijk feit vastgesteld over oversteken dat de uitwisselingen van chromosoomsegmenten die recombinatie van gekoppelde genen veroorzaken plaatsvinden in het vierstrengige of tetradstadium, in de profase van meiotische deling. Hier hebben de homologe chromosomen gepaard, is elke homoloog gesplitst in twee chromatiden bijeengehouden door onverdeelde centromeer en worden Chiasmata waargenomen in de tetrad.
Het wordt onthuld door zorgvuldige observatie dat bij elk chiasma slechts twee van de vier chromatiden kruisen terwijl de twee andere chromatiden hun oorspronkelijke continuïteit behouden. Genetisch betekent het dat van de vier chromosomen die uit meiose voortkomen, er maar twee op een bepaald niveau kunnen oversteken.
Recombinatie is een proces dat in verschillende situaties kan voorkomen, maar de meest populaire is in relatie tot meiose. Dit is dus een meiotisch proces dat een haploïde product genereert waarvan het genotype verschilt van de twee haploïde genotypen die het diploïde vormen. Het product van recombinatie is een recombinant.
Chiasma - type hypothese:
Morgan suggereerde dat de vorming van recombinanten kan worden verklaard door het optreden van crossing-over (dwz chiasma-vorming) tijdens meiose I. Dit leidde tot hypothese van het chiasma-type. De punten ter ondersteuning van deze hypothese zijn als volgt:
1. Tijdens profase I (di zygotene), homoloog chromosomenpaar (dwz synapsis), waarbij elk chromosoom uit twee chromatiden bestaat.
2. Tijdens pachytene van profase I breken twee niet-zusterchromatiden op homologe sites en komen dan weer samen.
3. Een dergelijke hereniging vindt plaats tussen niet-zuster-chromatiden en leidt tot fysieke uitwisseling.
4. Deze fysieke uitwisseling leidt tot herschikking van genen en dus vindt recombinatie plaats.
Synaptonemaal complex en het proces van oversteken:
Omdat het een vaststaand feit is dat DNA het materiaal van het gen is, wordt duidelijk dat oversteken gepaard gaat met het verbreken en opnieuw verbinden van DNA. Tijdens synapsis, lijnt een moleculaire scaffold die bekend staat als Synaptonemal complex de DNA-moleculen van de twee homologe chromosomen zij aan zij.
Met het resultaat kan de DNA-streng van één chromosoom, dat wil zeggen een chromatide, paren met de overeenkomstige DNA-streng van de ander, waar hun basensequentie overeenkomt. Breuk en nieuwe synthese van één DNA-streng met behulp van een streng van ander chromatide DNA als sjabloon resulteert in kruising.
Cross-over omvat het verbreken en weer verbinden van chromosomen in het Synaptonemal-complex, dat kan worden waargenomen met behulp van cytologische methoden. Er is echter geen kruising waargenomen bij mannelijke Drosophila tijdens spermavorming. Evenzo vindt er geen recombinatie van genen plaats tijdens meiose.