DNA-replicatie: opmerkingen over semi-conservatieve replicatie van DNA
Lees dit artikel voor meer informatie over de DNA-replicatie: opmerkingen over semi-conservatieve replicatie van DNA!
Replicatie is het proces van carbonkopieën. Hiervoor functioneert DNA als een eigen sjabloon. Daarom is DNA-replicatie een autokatalytische functie van DNA.

Afbeelding met dank aan: ehrig-privat.de/ueg/images/dna-replic.jpg
Het treedt meestal op tijdens de S-fase van de celcyclus wanneer de chromosomen in zeer uitgebreide vorm zijn. Zoals voorgesteld door Watson en Crick, is DNA-replicatie semiconservatief (een type replicatie waarbij één streng van de dochter-duplex is afgeleid van de ouder terwijl de andere streng opnieuw wordt gevormd).
Dit wordt uitgevoerd door de scheiding van twee strengen. De gescheiden strengen functioneren als sjablonen. De nieuwe strengen opgebouwd over de sjablonen van oude strengen hebben complementaire basenparen (A tegenover T en G tegenover C). De twee dochters-DNA-moleculen die aldus worden gevormd, zullen koolstof-kopieën van het oudermolecuul zijn, maar zullen één nieuwe streng en één oude streng hebben.
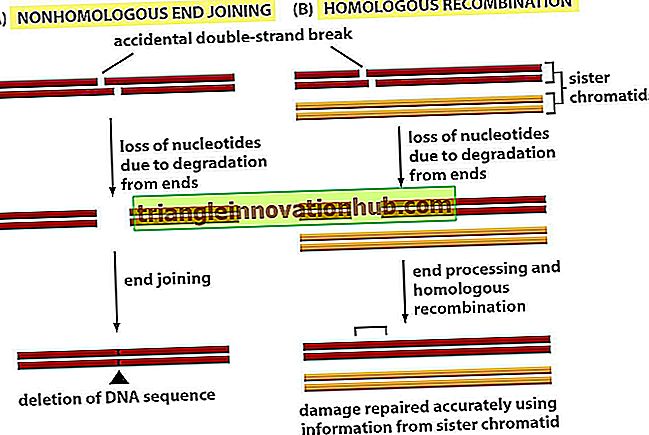
Taylor et al (1957) voerden delende cellen van worteluiteinden van tuinboon (Vicia faba) met radioactief 3H dat thymine bevat in plaats van normaal thymine. Thymine wordt ingebouwd in DNA dat het structurele element van chromosomen is. Taylor et al. Ontdekten dat alle chromosomen radioactief werden.
Gelabelde thymine werd vervolgens vervangen door de normale. De volgende generatie kreeg radioactiviteit in een van de twee chromatiden van elk chromosoom, terwijl de radioactiviteit van de volgende generatie aanwezig was in 50% van de chromosomen (figuur 6.9). Dit is alleen mogelijk als van de twee strengen van een chromosoom, de ene opnieuw wordt gevormd, terwijl de andere bij elke replicatie is geconserveerd, dit semiconservatieve replicatie is.

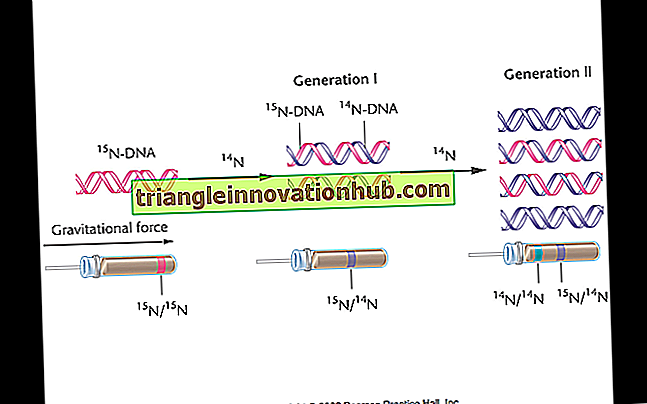
Semi-conservatieve replicatie van DNA werd bewezen door het werk van Mathew Meselson en Franklin Stahl (1958). Ze groeiden Escherichia coli gedurende vele generaties in een medium met zware isotoop van stikstof, in de vorm van 15 NH4C1, totdat het bacteriële DNA volledig werd gemerkt met zware isotoop.
De gemerkte bacteriën werden vervolgens naar vers medium met normale of 14 N stikstof verschoven. Monsters werden voor elke generatie genomen (één generatie duurt 20 minuten terwijl E. coli zich in 20 minuten verdeelt) en het DNA werd getest op de zware isotoop van stikstof door dichtheidsgradiëntcentrifugatie met behulp van cesiumchloride. Cesiumchloride is sterk in water oplosbaar zwaar zout.
Wanneer centrifuge met hoge snelheid wordt gecentrifugeerd (zeg 50.000 omwentelingen per minuut), vormt het zout een dichtheidsgradiënt met het zwaarste meest geconcentreerde gebied aan de onderkant en achtereenvolgens minder geconcentreerde lichtere naar het oppervlak. Wanneer DNA wordt gemengd met cesiumchloride, zal het op een bepaalde hoogte bezinken bij centrifugeren, zwaarder naar de basis en lichter naar boven (Fig. 6.10).
Fluorchroom, ethidemiumbromide genoemd, wordt gebruikt om het contrast te verbeteren, aangezien het fluorochroom specifiek is voor DNA. Meselson en Stahl vonden dat DNA van de eerste generatie hybride of intermediair was ( 15 N en 14 N). Het vestigde zich in cesiumchloride-oplossing op een niveau hoger dan het volledig gemerkte DNA van ouderbacteriën ( 15 N 15 N). De tweede generatie bacteriën na 40 minuten bevatte twee soorten DNA, 50% licht ( 14 N I4 N) en 50% tussenproduct ( 15 N I4 N).
De derde generatie bacteriën na 60 minuten bevatte twee soorten DNA, 25% tussenproduct ( 15 N 14 N) en 75% licht ( 14 N 14 N) in een verhouding van 1: 3. De vierde generatie bevatte na 80 minuten 12, 5% 15 N 14 N en 87, 5% 14 N 14 N DNA in een verhouding van 1: 7.
Deze waarneming is alleen mogelijk als de twee strengen DNA-duplex zich scheiden op het moment van replicatie en fungeren als een matrijs voor de synthese van nieuwe complementaire strengen van DNA met normaal of 14 N. Dit zal twee DNA-duplexen produceren met één oude streng ( 15 N) en één nieuwe streng ( 14 N).

Tijdens de vorming van de tweede generatie scheiden 15 N- en 14 N-strengen DNA-duplex zich af om als matrijzen te functioneren, zodat 50% van de nieuwe DNA-duplexen alleen normale of l4N- strengen bezitten, terwijl nog eens 50% zowel 15N- als 14N-strengen heeft (Fig. 6.11 en 6.12). Op deze manier wordt bij elke replicatie één streng ouder-DNA in de dochter geconserveerd terwijl de tweede vers gesynthetiseerd is.