Plasmamembraan: structuur en functies van plasmamembraan
Lees dit artikel voor meer informatie over de membraanstructuur, transportmodificatie en functies van plasmamembraan!
Plasma membraan:
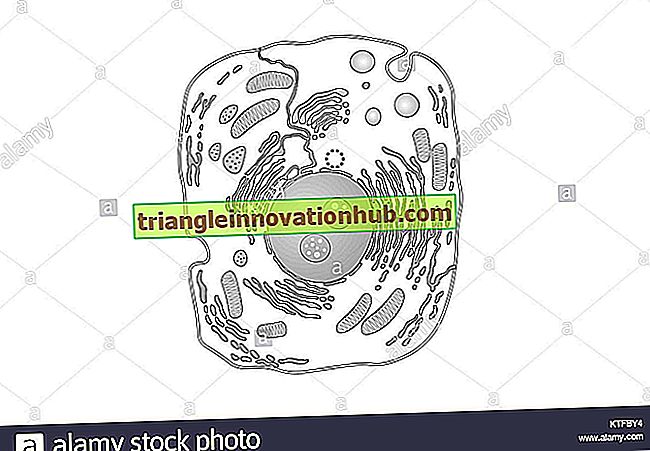
Plasmamembraan of plasma-lemma is een biomembraan dat aan de buitenkant van het cytoplasma voorkomt in zowel prokaryoten als eukaryote cellen.
Het scheidt het cellulaire protoplasma van zijn externe omgeving. Prokaryotische cellen hebben geen interne vliezige partities. De laatste komen voor in eukaryote cellen als bedekking van verschillende celorganellen zoals kern, mitochondria, plastiden, lysosomen, Golgi-lichaampjes, peroxisomen, enz.
Biomembranen vormen het endoplasmatisch reticulum. Ze komen ook voor op thylakoïden binnen plastiden of cristae in de mitochondriën. Vacuolen worden van cytoplasma gescheiden door een membraan genaamd tonoplast. Alle biomembranen zijn dynamisch van aard en vertonen voortdurend veranderingen in vorm, grootte, structuur en functie. Plasmamembraan werd ontdekt door Schwann (1838). Het werd genoemd als celmembraan door Nageli en Cramer (1855). Het membraan kreeg de naam van plasma-lemma van Plowe (1931).
Chemische aard van membranen:
Chemisch gezien bestaat een biomembraan uit lipiden (20-40%), eiwitten (59-75%) en koolhydraten (1-5%). De belangrijke lipiden van het membraan zijn fosfolipiden (ongeveer 100 soorten), sterolen (bijv. Cholesterol), glycolipiden, sphingolipiden (bijv. Sphingomyeline, cerebrosiden).
Koolhydraten die in het membraan aanwezig zijn, zijn vertakte of niet-vertakte oligosacchariden, bijvoorbeeld hexose, fucose, hexoamine, siaalzuur, enz. Eiwitten kunnen vezelig of bolvormig, structureel, drager, receptor of enzymatisch zijn. Ongeveer 30 soorten enzymen zijn opgenomen in verschillende biomembranen, bijv. Fosfatasen, ATP-ase esterasen, nucleasen, enz.
De lipidemoleculen zijn amfiatisch of amfipathisch, dat wil zeggen dat ze zowel polaire hydrofobe (waterminnende) als niet-polaire hydrofobe (waterafstotende) uiteinden bezitten. Het hydrofiele gebied heeft de vorm van een kop, terwijl het hydrofobe gedeelte twee staarten vetzuren bevat.
Hydrofobe staarten treden meestal op in de richting van het midden van het membraan. Eiwitmoleculen bezitten ook zowel polaire als niet-polaire zijketens. Gewoonlijk zijn hun polaire hydrofiele verbindingen naar de buitenzijde gericht. De niet-polaire of hydrofobe koppelingen worden ofwel binnengevouwen gehouden of gebruikt om verbindingen tot stand te brengen met het hydrofobe deel van de lipiden. Verschillende soorten modellen zijn naar voren gebracht om de structuur van een biomembraan te verklaren. Des te belangrijker zijn Lamellar en Mosaic.
Lamellar-modellen (= Sandwich-modellen):
Het zijn de vroege moleculaire modellen van biologische membranen. Volgens deze modellen wordt aangenomen dat biologische membranen een stabiele gelaagde structuur hebben.
Danielli en Davson Model:
Het eerste lamellaire model werd voorgesteld door James Danielli en Hugh Davson in 1935 op basis van hun fysiologische studies. Volgens Danielli en Davson bevat een biomembraan vier moleculaire lagen, twee van fosfolipiden en twee van eiwitten. Fosfolipiden vormen een dubbele laag.
De dubbellaag van fosfolipiden wordt aan beide zijden bedekt door een laag gehydrateerde globulaire of a-eiwitmoleculen. De hydrofiele polaire koppen van de fosfolipidemoleculen zijn gericht op de eiwitten. De twee worden bij elkaar gehouden door elektrostatische krachten. De hydrofobe niet-polaire staarten van de twee lipidelagen zijn gericht naar het centrum waar ze samen worden gehouden door hydrofobe bindingen en van der Waals-krachten.

Robertson Model:
J. David Robertson (1959) wijzigde het model van Danielli en Davson door voor te stellen dat de lipide dubbellaag op de twee oppervlakken wordt bedekt door verlengde of (3-eiwitmoleculen .Een verschil in de eiwitten van de buitenste en binnenste lagen werd ook voorgesteld, bijv. mucoprotine aan de buitenkant en niet-mucoïde eiwitten aan de binnenkant.
Robertson werkte op het plasmamembraan van rode bloedcellen onder elektronenmicroscoop. Hij gaf het concept eenheidsmembraan wat betekent dat:
(i) Alle cytoplasmatische membranen hebben een vergelijkbare structuur van drie lagen met een elektronen-transparante fosfolipide dubbellaag die met zand is bedekt tussen twee elektronen dichte lagen van eiwitten,
(ii) Alle biomembranen zijn ofwel gemaakt van een eenheidsmembraan of een veelvoud van eenheidsmembraan. Het eenheidsmembraan van Robertson wordt ook een sleepaminair membraan genoemd. Het heeft een dikte van ongeveer 75 A met een centrale lipidelaag van 35 A dik en twee perifere eiwitlagen van 20Aeach. Volgens Robertson, als een membraan meer dan drie lagen bevat, of dikker is dan 75A, moet het een veelvoud van eenheidsmembraan zijn.
Mozaïekmodel:
Vloeibaar mozaïek model. Het is het meest recente model van een biomembraan voorgesteld door Singer en Nicolson in 1972.
1. Volgens dit model heeft het membraan geen uniforme opstelling van lipiden en eiwitten maar is het in plaats daarvan een mozaïek van de twee. Verder is het membraan niet vast maar is het quasi-vloeibaar.
2. Het postuleert dat de lipidemoleculen aanwezig zijn in een viskeuze dubbellaag zoals in een lamellair model. Eiwitmoleculen komen op plaatsen zowel binnen als buiten de lipide dubbellaag voor. De interne eiwitten worden intrinsieke of integrale eiwitten genoemd, terwijl de externe eiwitten extrinsieke of perifere eiwitten zijn.
De integrale of intrinsieke eiwitten vormen 70% van de totale membraaneiwitten en gaan in de lipidedubbellaag in verschillende diepten. Sommigen van hen lopen door de lipide dubbellaag. Ze worden tunnelproteïnen genoemd die afzonderlijk of in een groep kanalen vormen voor de doorgang van water en wateroplosbare stoffen.
3. De eiwitten verschaffen de structurele en functionele specificiteit voor de membranen. Verder aangezien de lipidedubbellaag quasifluïde is, kunnen de membraaneiwitten lateraal verschuiven en vandaar flexibiliteit en dynamiek aan het membraan verschaffen.
Veel membraaneiwitten functioneren als enzymen, sommige gedragen zich als per-meases voor gefaciliteerde diffusie en een paar eiwitten fungeren als dragers omdat ze verschillende stoffen actief over het membraan transporteren. Bepaalde andere eiwitten functioneren als receptoren voor hormonen, herkenningscentra en antigenen. Een deel van het lipide aan het buitenoppervlak is gecomplexeerd met carbohydraten om glycolipiden of glycocalyx te vormen.

Modificaties van celmembraan:
1. Microvilli:
Het zijn vingerachtige eva-minaties van 0, 6-0, 8 μm lengte en 0, 1 μm diameter die gevonden worden op het vrije oppervlak van cellen die betrokken zijn bij absorptie, bijv. Darmcellen, hepatische cellen, mesotheelcellen, urineveuze tubuli. Het oppervlak met microvilli wordt gegroefde rand of borstelrand genoemd.
Microvilli vergroot het oppervlak meerdere keren. Ze worden ondersteund door een web van microfilamenten, actine samen met myosine, tropomysoïne, spectrine, enz. De nauwe ruimten tussen microvilli nemen deel aan pinocytose.

2. Mesosomes:
Het zijn plasmalemma-infouverties gevonden in bacteriën. Eén type mesosoom is intern gehecht aan de nucleoïde. Het is vereist voor nucleoid-replicatie en celdeling.

3. Junctional Complexes:
Het zijn contacten tussen naburige cellen die in het geval van dierlijke cellen worden gescheiden door ruimten van 150-200 Á gevuld met weefselvloeistof. De belangrijke zijn:
(i) Interdigitaties:
Er is een interlocking van vingerachtige membraanuitgroei tussen twee aangrenzende cellen. Interdigitaties vergroten het contactoppervlak tussen twee cellen voor uitwisseling van materialen.
(ii) Intercellulaire bruggen:
Projecties van aangrenzende cellen maken contact voor snelle geleiding van stimuli.
(iii) Strakke kruispunten:
(Zonulae Occludentes, singular- Zonula Occludens). Hier worden plasmamembranen van twee aangrenzende cellen op een reeks punten versmolten met een netwerk van richels of afdichtende strengen. Strakke juncties komen voor in epithelia met hoge elektrische weerstand en waar filtratie door de cellen moet plaatsvinden, bijv. Capillairen, hersencellen, het verzamelen van tubuli van nieren.

(iv) Gap Junctions:
De aangrenzende cellen hebben protoplasmatische verbindingen via speciale eiwitcilinders die connexons worden genoemd. Elke connexon is gemaakt van zes identieke eiwitsubeenheden rond een hydrofiel kanaal.

(v) Plasmodesmata:
Het zijn protoplasmatische bruggen tussen plantencellen die voorkomen in celmuurputten of poriën.
(vi) Desmosomen:
(Maculae Adherentes, enkelvoud-Macula Adherens). Aangrenzende membranen bezitten schijfvormige verdikkingen van ongeveer 0, 5 (am diameter, een aantal tonofibrillen (= tonofilamenten) en trans-membraan linkers ingebed in dicht intercellulair materiaal. Desmosomen fungeren als puntlassen en worden daarom vlek desmosomen genoemd Ze komen voor in epithelia onderworpen tot verstoring.

(vii) Terminal Bars:
(Belt Desmosomes, Zonulae Adherentes, singular-Zonula Adherens. Intermediary Junction). Eindstaven zijn desmosomen zonder tonofibrillen. Banden van verdikkingen komen voor op het binnenoppervlak van het membraan. De banden bevatten microfilamenten en intermediaire filamenten.
Functies van celmembranen:
1. De belangrijkste functie van celmembranen is compartimentering. Als plasmamembranen scheiden ze de cellen van hun externe omgeving. Als organelbekledingen laten ze de celorganellen toe om hun identiteit, specifieke interne omgeving en functionele individualiteit te behouden.
2. De membranen laten de stroom van materialen en informatie tussen verschillende organellen van dezelfde cel evenals tussen één cel en een andere toe.
3. Als plasmodesmata en spleetovergangen verschaffen de biomembranen organische verbindingen tussen naburige cellen.
4. Plasmamembranen evenals andere membranen van de organellen hebben selectieve permeabiliteit, dat wil zeggen ze laten alleen geselecteerde stoffen naar binnen doorgaan naar geselecteerde graden. De membranen zijn ondoordringbaar voor anderen.
5. Biomembranen hebben de eigenschap van remanentie, dat wil zeggen dat ze de doorgang van stoffen die al toegelaten zijn niet toelaten.
6. Plasmamembraan bezit specifieke stoffen aan het oppervlak die functioneren als herkenningscentra en bevestigingspunten.
7. Stoffen die aan het celmembraan zijn bevestigd, bepalen antigeenspecificiteit. Glycoforines die aanwezig zijn op het oppervlak van erytrocyten functioneren als antigeendeterminanten. Histocompatibiliteit-antigenen duiden aan of een vreemde cel of weefsel moet worden opgenomen of afgewezen.
8. Celmembraan heeft receptoren voor bepaalde hormonen. Het hormoon combineert met zijn specifieke receptoren en verandert ofwel membraanpermeabiliteit of activeert enzym adenylaatcyclase om cyclisch AMP uit ATP te produceren. cAMP activeert vervolgens een reeks enzymen om een bepaalde functie uit te voeren.
9. Membranen hebben dragereiwitten voor actief transport.
10. Celmembranen bevatten enzymen voor het uitvoeren van bepaalde reacties op hun oppervlak, bijvoorbeeld ATP-ase (voor ATP-synthese en afgifte van energie uit ATP), fosfatasen, esterasen, enz.
11. Bepaalde celmembranen (bijv. Plasmamembraan in bacteriën, thylakoïde membranen van chloroplasten, binnenmi- croondriaal membraan) bezitten elektronentransportsystemen.
12. Membraaninvouwingen worden gebruikt voor bulkopname van materialen door endocytose.
Membraan transport:
Passage van stoffen over biologische membranen of celmembranen wordt veroorzaakt door de volgende methoden:
A. Transport van water:
(I) Osmose:
Osmose is de diffusie van water of oplosmiddelmoleculen door plasmamembraan van lage osmotische druk tot hoge osmotische druk, dat wil zeggen van hoog watergehalte naar laag watergehalte. Plasmamembraan werkt als een differentieel membraan dat de beweging van watermoleculen in en uit het behoud van de metabolieten mogelijk maakt.
B. Transport van ionen en kleine moleculen:
(II) Passief vervoer:
Het is een modus van membraantransport waarbij de cel geen energie uitgeeft en geen speciale activiteit vertoont. Het transport verloopt volgens concentratiegradiënt. Het is van twee soorten, passieve diffusie en gefaciliteerde diffusie.
(a) Passieve diffusie of transport over celmembranen:
Hier speelt het celmembraan een passieve rol bij het transport van stoffen eroverheen. Passieve diffusie kan plaatsvinden via lipidenmatrix van het membraan of met behulp van kanalen.
(i) Lipide oplosbare stoffen:
Het werd ontdekt door Overton (1900) dat lipide oplosbare stoffen snel door het celmembraan gaan volgens hun concentratiegradiënt. Op basis van deze bevinding stelde Overton voor celmembranen van lipiden te maken.
(ii) Kanaaltransport:
Membraan heeft kanalen in de vorm van tunnelproteïnen die geen lading dragen. Ze laten water en oplosbare gassen (CO 2 en O 2 ) door in overeenstemming met hun concentratiegradiënt. Osmose is een voorbeeld van zo'n transport.
Als twee oplossingen van verschillende concentraties worden gescheiden door een semi-permeabel membraan, bewegen de oplosmiddelmoleculen over het membraan van minder geconcentreerd naar de meer geconcentreerde oplossing. Dit proces - de diffusie van oplosmiddelmoleculen in een regio waarin zich een hogere concentratie opgeloste stof bevindt waarvoor het membraan ondoordringbaar is, wordt osmose genoemd.
Filtratie is diffusie onder druk over een membraan met minieme poriën. Ultrafiltratie vindt plaats tijdens glomerulaire filtratie in de nieren. Dialyse is het proces van het scheiden van kleine deeltjes (bijv. Kristallijne opgeloste stoffen) van grotere deeltjes (bijv. Colloïden) vanwege verschil in de snelheid van diffusie over een membraan met zeer kleine poriën.
(b) Gefaciliteerde verspreiding:
Het gebeurt door het gebruik van speciale membraaneiwitten die permeases worden genoemd. Wanneer dergelijk door transport gemedieerd transport uit een gebied van grotere concentratie is, is energie niet vereist en wordt het proces gefaciliteerde diffusie genoemd. Als gevolg hiervan is de transportsnelheid stereospecifiek.

Het invoeren van glucose in rode bloedlichaampjes is een gefaciliteerde diffusie.
Het proces van gefaciliteerde diffusie omvat de volgende stappen:
1. Diffuserende moleculen combineren met de specifieke dragereiwitmoleculen die drager-eiwitcomplexen vormen.
2. De vorm van dragereiwitmolecuul verandert in reactie op het diffunderende molecuul, zodat de membraan-gebonden dragereiwitcomplexen kanalen vormen.
3. De vorm van het dragereiwitmolecuul verandert in reactie op het diffuserende molecuul, waardoor het molecuul het plasmamembraan kan passeren.
4. Zodra het diffunderende molecuul de andere kant heeft bereikt, verlaagt de verandering in vorm van het dragermolecuul (conformationele verandering) zijn affiniteit met het diffunderende molecuul en maakt het mogelijk dat het wordt vrijgegeven.
5. Na het vrijkomen van het diffunderende molecuul hervat het dragereiwitmolecuul de oorspronkelijke vorm.
De gefaciliteerde diffusie maakt het mogelijk dat moleculen het anders ondoordringbare of slecht doordringbare membraan passeren.
Gefaciliteerde diffusie verschilt van eenvoudige diffusie in de volgende functies:
(i) Gefaciliteerde diffusie is stereospecifiek (L- of D-isomeer wordt getransporteerd).
(ii) Het toont verzadigingskinetiek.
(iii) Gefaciliteerde diffusie vereist een drager voor transport door het membraan. De dragereiwitmoleculen bewegen heen en weer over het membraan door thermische diffusie.
(III) Actief vervoer:
Het is bergopwaartse beweging van materialen over de membranen waar de opgeloste deeltjes bewegen tegen hun chemische concentratie of elektrochemische gradiënt. Deze vorm van transport vereist energie die bijna uitsluitend wordt verschaft door hydrolyse van ATP.
Actief transport vindt plaats in het geval van zowel ionen als niet-elektrolyten, bijv. Zoutopname door plantencellen, ionen, glucose en fenolftaleïne in het geval van niertubuli, natrium en kalium in het geval van zenuwcellen, enz. Het wordt ondersteund door verschillende bewijzen:
(a) De absorptie wordt verminderd of gestopt met de afname van het zuurstofgehalte in de omringende omgeving.
(b) Metabole remmers zoals cyaniden remmen de absorptie.
(c) Cellen verzamelen vaak zouten en andere stoffen tegen hun concentratiegradiënt.
(d) Actief transport toont verzadigingskinetiek, dat wil zeggen, de snelheid van transport neemt toe met toename van de opgeloste concentratie tot een maximum wordt bereikt. Buiten deze waarde neemt de snelheid van het membraantransport niet toe, wat aangeeft dat het plaatsvindt door het gebruik van speciale organische moleculen, dragermoleculen, dragerdeeltjes of dragereiwitten.
De dragermoleculen zijn ATP-assen, enzymen die de hydrolyse van ATP katalyseren. De belangrijkste van deze ATPasen is Na + -K + ATP ase die ook bekend staat als de Na + -K + pomp. Er zijn bovendien H + -K + ATPasen in het maagslijmvlies en de niertubuli.
Er is een speciaal dragermolecuul voor elk opgeloste deeltje. De drager heeft zijn bindingsplaats op twee oppervlakken van het membraan. De opgeloste deeltjes worden gecombineerd met de drager om een drager-opgelost complex te vormen. In de gebonden toestand ondergaat de drager een conformatieverandering die de opgeloste stof naar de andere kant van het membraan transporteert. Energie wordt gebruikt om de conformatieverandering in de drager tot stand te brengen. Het wordt geleverd door ATP. In het proces wordt ATP gedefosforyleerd om ADP te vormen. Carrier-eiwitten zijn van drie soorten.
1. Uniport:
Ze vervoeren slechts één stof.
2. Symports:
In sommige gevallen vereist transport de binding van meer dan één stof aan het transporteiwit en worden de stoffen samen over het membraan getransporteerd. Een voorbeeld is de symport in het darmslijmvlies die verantwoordelijk is voor het co-transport door gefaciliteerde diffusie van Na + en glucose uit het intestinale lumen in mucosale cellen.
3. Antipoorten:
Ze wisselen de ene substantie in voor de andere. De Na + -K + ATPase is een typische antipoort.

Veel dierlijke cellen hebben een natrium-kalium-uitwisselingspomp op hun plasmamembraan. Een soortgelijke protonpomp werkt in chloroplasten, mitochondriën en bacteriën Na + -K + uitwisselingspomp werkt met behulp van enzym ATP-ase dat ook fungeert als een dragermolecuul.

Het enzym hydrolyseert ATP om energie vrij te maken. De energie wordt gebruikt om conformationele veranderingen in de draaggolf teweeg te brengen. Voor elk gehydrolyseerd ATP-molecuul worden drie Na + -ionen naar buiten gepompt en twee K + -ionen worden naar binnen gepompt.
Na + - K + wisselpomp voert de volgende functies uit: (i) Behoudt een positieve potentiaal aan de buitenkant van het membraan en relatief elektronegatief potentieel aan de binnenkant,
(ii) De pomp creëert een rustpotentiaal in de zenuwcellen,
(iii) De pomp handhaaft de waterhuishouding van levende cellen.
(iv) Het helpt bij de vorming van urine,
(v) Het neemt deel aan de uitscheiding van zout zoals bij zeedieren. Zeemeeuwen en pinguïns drinken zeewater. Ze scheiden overtollig zout af via de neusklieren. De nasale zoutklieren hebben een natrium-kaliumpomp in de plasmamembranen van hun cellen. Na + -ionen worden actief gepompt. Chloorionen passeren passief. De neussecretie van de twee vogels bezit 1, 5 - 3, 0 maal meer NaCl-concentratie dan die aanwezig is in het bloed.
(vi) De niet-afgescheiden en niet-gemetaboliseerde overmatige Na + -ionen die aanwezig zijn in het extracellulaire fluïdum hebben de neiging terug te gaan in de cellen. Andere stoffen worden gecombineerd met natriumionen en passeren ze naar binnen, bijvoorbeeld glucose, aminozuren in de darm. Het fenomeen wordt secundair actief transport genoemd in vergelijking met Na + -K + -wisselaarspompen, dat primair actief transport wordt genoemd.

Andere belangrijke pompen zijn onder meer Calciumpomp (RBC's, spieren), K + pomp, CP pomp, K + -H + wisselpompen. De laatste komt voor in wachtcellen.
Actief transport is een middel om (i) de meeste voedingsstoffen uit de darm te absorberen (ii) reabsorptie van bruikbaar materiaal uit de urinevormende tubuli (iii) snelle en selectieve absorptie van nutriënten door cellen (iv) behoud van een membraanpotentiaal (v) onderhoud van rustpotentieel in zenuwcellen (vi) behoud van water en ionische balans tussen cellen en extracellulaire vloeistof, (vii) uitscheiding van zoute klieren.
C. Transport van vaste deeltjes (bulktransport):
Bulktransport zowel naar binnen als naar buiten vindt plaats over het plasmamembraan door invaginatie en verdamping van het membraan. Bulktransport is nuttig bij het dragen van grote moleculen die het moeilijk zouden hebben om normaal door het celmembraan te gaan. Endocytose en exocytose zijn de twee manieren waarop bulktransport wordt bereikt.
(IV) Endocytose is het proces waarbij grote deeltjes van voedingsstoffen of vreemde stoffen worden overspoeld. Afhankelijk van de aard van de stoffen kan de endocytose zijn:
(i) Pinocytose of het drinken van cellen is het proces van inname van vloeibaar materiaal door de cel.
(ii) Micro-pinocytose is de pinocytose van subcellulair of sub-microscopisch niveau.
(iii) Rhophaeocytose is de overdracht van kleine hoeveelheden cytoplasma samen met hun opname.
(iv) Fagocytose versmelt grote deeltjes vast voedsel of vaste materie door de cel.
(V) Exocytose is het exsudatieproces van de secretaire producten naar buiten het cytoplasma van de cel. Het is ook bekend als emeicytosis of cel braken. In cellen van de pancreas bewegen de vacuolen die enzymen bevatten van het binnenste van het cytoplasma naar het oppervlak. Hier versmelten ze met plasmamembraan en ontladen ze hun inhoud naar buiten.