Golgi Complex: terminologie, voorkomen, morfologie, structuur en andere details
Golgi Complex: terminologie, voorkomen, morfologie, structuur, structurele en biochemische polarisatie, chemische samenstelling, functies en oorsprong!
In 1898 ontdekte Golgi met behulp van een zilverkleuring een reticulaire structuur in het cytoplasma. De naam "Golgi-apparaat" die in het algemeen aan deze structuur wordt gegeven, is verwarrend omdat het een duidelijke relatie suggereert met de fysiologische processen van de cel.
Tegenwoordig lijkt het geschikter om de naam "Golgi-substantie" of "Golgi-complex" te gebruiken om naar dit materiaal te verwijzen dat speciale kleuringseigenschappen heeft. Omdat de brekingsindex vergelijkbaar is met die van de matrix, is het Golgi-complex moeilijk waar te nemen in levende cellen. Het gebruik van de elektronenmicroscoop heeft gezorgd voor een duidelijk beeld van deze component en de submicroscopische structuur is onthuld.
Jarenlang werd gedacht dat het Golgi-complex een artefact was van verschillende fixatie- en kleuringsprocedures. Met andere woorden, veel wetenschappers geloofden dat de structuur die werd waargenomen tijdens talloze microscopieprocedures en de Golgi werd genoemd, in werkelijkheid niet bestond in de levende cel. Guilliermond (1923), Parat (1927), Walker en A lien (1921), ontstonden twijfels over het bestaan van het Golgi-complex.
Terminologie van Golgi-complex:
Holmgren verwees naar het Golgi-complex als trophospongium (Cajal noemde het de Golgi-Holmgren-grachten). Baker gebruikte de term lipochondria vanwege de veronderstelde lipideninhoud. De term Dalton Complex werd gegeven naar de naam van zijn waarnemer Dalton in 1952. Sjostrand stelde de term cyto-membranen voor voor het Golgi-systeem.
Sosa heeft de volgende nomenclatuur voor het Golgi-complex voorgesteld:
1. Golgiokinesis :
Indeling van het Golgi-apparaat tijdens nucleaire splitsing.
2. Golgiosomes :
Bloedlichaampjes geproduceerd door de Golgiogenesis worden Golgiosomes genoemd, die worden beschreven als Golgi-materiaal in ongewervelde dieren.
3. Golgiolyse :
Werkwijze voor het oplossen van het Golgi-apparaat.
4. Gogiorrhexis :
Fragmentatie op het Golgi-apparaat.
5. Golegiogenesis :
Vorming en differentiatie van het Golgi-lichaam tijdens de embryonale ontwikkeling.
6. Golgio-cytoarchitecture :
Studie van de structuur van de cel in relatie tot Golgi-apparatuur.
Voorkomen:
Het Golgi-complex komt voor in alle cellen behalve de prokaryotische cellen (namelijk Mycoplasma, bacteriën en blauwgroene algen) en eukaryote cellen van bepaalde schimmels, spermacellen van bryofyten en pteridofyten, cellen van rijpe zeefbuizen van planten en volgroeide zaadcellen en rood bloed. cellen van dieren.
Morfologie van Golgi-complex:
De morfologie van het Golgi-complex varieert van cel tot cel, afhankelijk van het type cel waarin ze worden gevonden. Twee vormen van Golgi-complex zijn waargenomen.

1. Gelokaliseerd formulier :
In gepolariseerde cellen van gewervelde dieren (die basis en apex hebben), vindt het Golgi-complex afzonderlijk plaats en neemt het een vaste positie in. Het ligt tussen de nucleus en de uitscheidingspool. Dit kan het beste worden gezien in schildkliercellen, in exocriene cellen van de pancreas en de slijmcellen van de darm.
2. Diffuse vorm :
In sommige gespecialiseerde cellen van gewervelde dieren (zenuwcellen en levercellen), worden in de meeste plantencellen en in de cellen van ongewervelde dieren verschillende eenheden van het Golgi-complex verspreid aangetroffen samen met de elementen van endoplasmatisch reticulum. Elke eenheid wordt een dictyosome genoemd. In levercellen komen er maar liefst 50 dictyosomen per cel voor en in bepaalde plantencellen kan hun aantal oplopen tot honderden.
Vorm :
De vorm van het Golgi-complex is vrij variabel in verschillende somatische celtypen van dieren. Zelfs in dezelfde cel zijn er variaties in verschillende functionele fasen. De vorm is echter constant bij elk celtype. Het varieert in vorm van een compacte massa tot een verspreid, filamenteus netwerk.
Aantal:
Het aantal Golgi-stapels per cel varieert enorm, afhankelijk van het celtype, van slechts een tot honderden. Er is een enkele grote in sommige cellen terwijl in het geval van Paramoeba er twee zijn. In Stereomyxa (een soort van Amoeba) zijn er veel Golgi-complexen. Zenuwcellen, levercellen en de meeste plantencellen hebben ook meerdere Golgi-complexen, ongeveer 50 in levercellen.
In algenrizoidachtigen wordt meer dan 25.000 Golgi-complex aangetroffen (Sievers 1965). Het Golgi-complex kan zelfs een groot deel van het celvolume in sommige gespecialiseerde cellen verklaren. Een voorbeeld is de slijmbekercel van het intestinale epitheel, die slijm in de darm afscheidt; de glycoproteïnen in mucus zijn in hoofdzaak geglycosyleerd in het Golgi-complex.
Grootte:
De grootte is eveneens variabel. Het is groot in de zenuw- en klierencellen en klein in de spiercellen. In het algemeen is het Golgi-complex goed ontwikkeld terwijl de cel in actieve toestand is. Wanneer de cel oud wordt, neemt het complex progressief af in grootte en verdwijnt het.
Positie:
De positie van het Golgi-complex is relatief vastgesteld voor elk celtype. In de cellen van ectodermale oorsprong is het Golgi-complex gepolariseerd vanaf de tijd van de embryonale toestand tussen de kern en de periferie (Cajal, 1914). In de secretoire exocriene cellen die in het algemeen een typische polarisatie hebben, wordt het Golgi-complex gevonden tussen de kern en de secretorische pool.
In de endocriene klieren is de polariteit van deze organoïde variabel, behalve in de schildklier, waar deze naar het midden van de follikel is gericht. In de jongere cellen en vaak in de oudere cellen ligt het meestal aan één kant van de kern, maar in bepaalde gevallen kan het volledig omringen. In ganglioncellen van de muis is de positie perinucleair.
Gedetailleerde structuur van Golgi Complex:
Dalton en Felix (1954) beschreven het Golgi-complex in de epididymis van de rat na het nemen van de eerste elektronenmicrofoto's.
De volgende beschrijving van het Golgi-complex is samengesteld op basis van het werk van verschillende auteurs:
1. Cisternae :
De cisternae of saccules zijn vergelijkbaar met het gladde oppervlak ER en verschijnen in doorsnede als stapels dicht op elkaar liggende door membranen begrensde zakjes. Het aantal saccules varieert van 4 tot 8 in de meeste celtypen van dieren en planten. In Euglena kan het aantal oplopen tot 20.
Het membraan van de saccules heeft een dikte van ongeveer 60 tot 70 A °, die een holte insluit ongeveer 150 A ° breed waarvan de randen vaak zijn verwijd. Volgens de meeste auteurs zijn er twee goed gedefinieerde gezichten van de cisternae dwz convex en concaaf; de laatste wordt algemeen aangeduid als het volwassen of vormende of distale vlak en de convexe zijde wordt verondersteld het onrijpe of excite of proximale vlak te zijn, de cisterane liggen in parallel array zijn van elkaar gescheiden door een ruimte van ongeveer 200 tot 300 A 0 .
Wat ze bij elkaar houdt is nog niet bekend, maar in enkele cellen wordt een dunne laag ondoorzichtig, soms dicht materiaal gezien tussen de saccules die op bepaalde gebieden prominenter zijn, waarop Amos en Grimstone (1968) de term knooppunten hebben toegepast. Mollenhauer et al., (1973) verkenden in zekere mate intercisternale elementen en plaques in een bepaald Golgi-complex.

2. Tubules :
Vanuit het perifere gebied van cisternae ontstaat een complex, anastomoserend vlak netwerk van tubuli met een diameter van 300 tot 500 A °. Clowes en Juniper (1969) hebben dit buisvormige netwerk vergeleken met een schijf van kant.
3. Vesicles :
De vesicles zijn kleine druppelachtige zakjes die aan de rand van de cisternae aan tubuli blijven vastzitten.
Ze zijn van de volgende twee typen:
(a) Gladde blaasjes :
De gladde vesicles hebben een diameter van 20 tot 80μ. Ze bevatten secretiemateriaal (wordt zo vaak secretoire blaasjes genoemd) en zijn afgetopt van de uiteinden van cisternale tubuli in het net. Vaak meer dan één tubuleaansluiting, en vermoedelijk te vullen, een enkele vormblaasje.
(b) Gecoate blaasjes :
De gecoate vesicles zijn bolvormige uitsteeksels, ongeveer 50 μm in diameter en met een ruw oppervlak. Ze worden gevonden aan de periferie van het organel, meestal aan de uiteinden van enkele tubuli en zijn morfologisch heel verschillend van de secretoire vesicles. Hun functie is onbekend.
4. Golgiaanse vacuoles :
Dit zijn grote afgeronde klodders die op het volwassen gezicht van Golgi aanwezig zijn. Deze worden gevormd door de uitgezette cisternae of door de fusie van uitscheidende blaasjes. De vacuolen zijn gevuld met een of andere amorfe of korrelige substantie.
Het Golgi-complex is structureel en biochemisch gepolariseerd:
Het Golgi-complex heeft twee verschillende gezichten: een cis, of een vormvlak en een trans- of volwassen gezicht. Het cis-oppervlak is nauw verbonden met een glad overgangsgedeelte van de ruwe ER. In secretiecellen is het trans-vlak het gezicht dat zich het dichtst bij het plasmamembraan bevindt: hier worden de grote afscheidingsblaasjes uitsluitend gevonden in associatie met het trans-oppervlak van een Golgi-stapel en het membraan van een zich vormend secretoir blaasje is vaak continu met dat van het trans-vlak van de laatste ("trans-meest") cisterna.
In tegenstelling daarmee zijn de kleine Golgi-vesicles gelijkmatiger gelokaliseerd langs de stapel, wordt er algemeen aangenomen dat eiwitten een Golgi-stapel binnengaan vanuit de ER aan de cis-zijde en om af te sluiten voor meerdere bestemmingen aan de trans-zijde; Noch hun exacte pad door het Golgi-complex, noch hoe ze van cisterna naar cisterna langs elke stapel reizen, zijn echter bekend.
De twee vlakken van het Golgi-complex zijn biochemisch verschillend. Een variatie in de dikte van de Golgi-membranen kan bijvoorbeeld in bepaalde gevallen over de stapel worden gedetecteerd, waarbij die aan de cis-zijde dunner zijn (ER-achtig) en die aan de trans-zijde dikker zijn (plasmamembraan-achtig).
Meer opvallend zijn de resultaten die worden verkregen wanneer bepaalde histochemische tests worden gebruikt in combinatie met elektronenmicroscopie om bepaalde eiwitten in het Golgi-complex te lokaliseren. Sommige van deze tests onthullen membraangebonden enzym, activiteiten die een duidelijke polariteit vertonen in hun lokalisatie binnen de Golgi-stack.
Een bijzonder intrigerende biochemische vondst was de ontdekking dat lysosomale enzymen, zoals zuurfosfatase, geconcentreerd zijn met de trans-meest cisterna van de Golgi-stapel en in enkele nabijgelegen beklede vesicles. Dit suggereert dat specifieke vesikels die naar lysosomen vertrekken, in dit gebied worden geassembleerd.
Uitscheidende eiwitten worden aangetroffen door histochemische methoden in alle gestapelde cisternae, hoewel de grote uitscheidende blaasjes waarin deze producten zijn geconcentreerd en alleen zijn geassocieerd met de trans-meest Golgi cisterna.
Chemische samenstelling:
Wat de chemische samenstelling van het Golgi-complex betreft, is aangetoond dat de volgende stoffen aanwezig zijn:
1. Fosfolipiden :
De samenstelling van fosfolipiden van Golgi-membranen ligt tussen die van endoplasmatische membranen en plasmamembranen.
2. Eiwitten en enzymen :
Golgi-complex van verschillende planten- en dierencellen vertonen grote variaties in de eiwit- en enzyminhoud. Enkele van de enzymen zijn ADPase, ATPase, NADPH cytochroom-C-reductase, glycosyl transferases, galactosyl transferase, thiamine pyrophosphate etc.
3. Koolhydraten :
Zowel planten- als dierlijke cellen hebben enkele gebruikelijke koolhydraatcomponenten, zoals glucosarine, galactose, glucose, mannose en fructose. Plant Golgi missen siaalzuur, maar het komt in hoge concentraties voor in de lever van de rat. Sommige koolhydraten zoals xylulose en arabinose zijn alleen in plantencellen aanwezig.
4. Vitamine С :
De fractie van vitamine С opgeslagen in het Golgi-complex is aangetoond door Tomitte. Volgens hem slaat Golgi complex vitamine С op en maakt het langzaam in het cytoplasma vrij in voldoende hoeveelheid om oxidatie van de celproducten te voorkomen.

Functies van Golgi-complex:
1. Vorming van acrosoom tijdens spermiogenese :
Tijdens de rijping van sperma speelt het Golgi-complex een rol bij de vorming van acrosoom (Burgos en Fawcett, 1955).

In vroege stadia verschijnt de Golgi als een bolvormig lichaam, bestaande uit cisterna's gerangschikt in parallelle stapels en talrijke kleine blaasjes. De later afgeknepen altijd van de cisternae. Naarmate de ontwikkeling vordert, wordt het Golgi-complex onregelmatig van vorm en worden grote vaculoles gevormd door dilataties van cisternale zakken.
In het midden van deze grote vacuole of vacuolen bevindt zich een dichte granule, de proacrosomale granule. Deze korrel die is afgeleid van het Golgi-complex blijft binnen de vacuole groeien door een proces dat bekend staat als aangroei. Deze vacuole en granule nadert de anterieure pool van het kernmembraan en vormt een acrosomale granule.
Met de verlenging van de spermatid spreidt het acrosomale vesikel zich over het nucleaire oppervlak uit en tenslotte bezwijkt het met het kernmembraan, waardoor het dopmateriaal wordt gevormd. De acrosomale granule wordt het acrosoom dat aan de top van de kern ligt en blijkbaar bepaalde enzymen omvat die bij het bevruchtingsproces betrokken zijn.
2. Synthese en secretie van polysacchariden:
Onderzoeken naar slijmbekercellen met autoradiografie en elektronenmicroscopie hebben het verband aangetoond tussen eiwitsynthese, koolhydraattoevoeging en sulfatering. De slijmbekercellen van de dikke darm produceren mucigen. Dit secretoire materiaal bevat een groot aandeel koolhydraten.
Het Golgi-complex bevindt zich net boven de kern. Naar het vrije oppervlak van de cel worden geleidelijk mucigen-korrels vergroot. De proximale cisternae van het Golgi-complex vertonen geen zwelling, maar op enige afstand over de stapel worden de distale cisternae vrij plotseling omgezet in mucigen korrels. De distale cisternae zetten om de 2-4 minuten continu om in mucigeenkorrels. Nieuwe proximale cisternae worden ter compensatie gevormd.
3. Rol in afscheiding:
Golgi-complex wordt geacht enige rol te spelen in de secretoire functie van een cel. Maar de vraag is dat ze sommige stoffen zelf uitscheiden of synthetiseren of dat ze gewoon een opslaghuis zijn waarin de secretoire producten die ergens anders in de cel worden uitgescheiden, gewoon worden opgeslagen en geconcentreerd.


Uit de studies van Palade et al. 1962 deze secretoire cyclus is nu goed gedefinieerd en omvat vier stappen in het geval van acinaire cellen van de alvleesklier en ze zijn:
(i) De opname van aminozuren in eiwitten aan het oppervlak van ruw endoplasmatisch reticulum.
(ii) Overdracht van deze ontluikende secretie-eiwitten in de cisternae van ruw endoplasmatisch reticulum.
(iii) Het intracellulaire transport van deze eiwitten naar het Golgi-complex.
(iv) De migratie van zymogeenkorrels naar de top van de cel waar ze in lumen werden geloosd.
4. Rol van het lichaam van Golgi in oögenese :
Srivastava (1965) heeft een kort overzicht gegeven van het Golgi-complex tijdens de oogenese. Volgens Afzelius (1956) bestaat het Golgi-complex van een zee-ei-ei, zoals te zien onder een elektronenmicroscoop, uit stapels lamellen vormende wanden van platte buidels, die af en toe opgezwollen kunnen zijn.
Er zijn enkele aanwijzingen voor transversale indelingen van deze lichamen. Sotelo (1959) en Sotelo en Porter (1959) hebben het Golgi-complex in rattenovum beschreven onder de elektronenmicroscoop en juxtra nucleaire lokalisatie van dit organel in vroege oöcyten gevonden.
In de volgende fase lossen deze op in fragmenten en in de derde fase deze bewegingen naar de cortex. In al deze gevallen blijft hun structuur bestaan uit dicht opeengepakte reeksen dunne, dubbele profielen (afgeplatte zakjes) en bolvormige blaasjes.
In de vroege oöcyten is het complex compact georganiseerd. In latere stadia worden discrete bundels van profielen, omgeven door kleine blaasjes, gevonden verspreid in de corticale zone. In de vroege oöcyten zijn het Golgi-complex en het centrosoom nauw met elkaar verbonden.
5. Absorptie van verbindingen :
Hirsch et al. Hebben ontdekt dat wanneer ijzersuiker aan een dier wordt gevoerd, ijzer wordt geabsorbeerd op het Golgi-complex (Kedrowsky). Van Teel heeft aangetoond dat Golgi-systemen ook verbindingen van koper en goud absorberen. Kedrowsky heeft aangetoond dat Golgi-complex van Opalina bismutose (verbinding van albumine en bismut) en protargol (verbinding van albumine en zilver) kan absorberen. Kirkman en Severinghaus stellen dus dat het Golgi-complex fungeert als een condensatiemembraan voor de concentratie van producten tot druppels of korrels.
6. Vorming van plantencelwanden :
De celwand van planten bestaat uit fibrillen die overwegend polysacchariden bevatten, samen met enkele lipiden en eiwitten. Tijdens cytokinese wordt een celplaat gevormd tussen de twee dochterkernen en eromheen een membraan dat later het plasmamembraan van de dochtercellen wordt. Er is duidelijk bewijs dat de polysacchariden worden gevormd in het Golgi-complex en worden overgebracht naar de nieuwe celwand die wordt neergelegd terwijl de cellen nog steeds groeien.

Stoffen zoals pectines en hemicelluloses, die de matrix vormen van de celplaat die de plasmamembranen scheidt, worden ook bijgedragen door het Golgi-complex.
7. Vorming van intracellulaire kristallen :
In de mariene isopode, Limnoria lingmorum, die een ingravende vorm is, zijn er aanwezige midglands waarvan de cellen uit kristallen bestaan. Deze variëren tot 30 A ° in lengte en 15 A ° dik. Het is bewezen dat deze kristallen worden gevormd door het Golgi-complex en waarvan bekend is dat ze eiwit en ijzer bevatten. Ze hebben geen omhullend membraan en zijn meestal bolvormig van vorm. Ze houden zich bezig met de secretoire activiteit.
8. Melkproteïne-druppelvorming :
In de melkgevende borstklier van muizen worden eiwitdruppeltjes geproduceerd die verwant zijn aan het Golgi-complex. Deze druppeltjes gaan gewoonlijk open op het celoppervlak door de fusie van hun omhullende membraan met het plasmamembraan.
9. Vorming van lysosomen en vacuolen :
Primaire lysosomen worden gevormd uit de Golgi-membranen op dezelfde manier als de afscheidende blaasjes. Er is goed bewijs dat dictyosomen hydrolytische enzymen accumuleren in hun meer volwassen gebieden. Sommige vacuolen in plantencellen bleken kleine hoeveelheden hydrolytische enzymen te bevatten en deze worden verondersteld afgeleid te zijn van het Golgi-complex.
10. Pigmentvorming:
In veel zoogdiertumoren en kankercellen is het Golgi-complex beschreven als de plaats van oorsprong van pigmentgranulaat (melanine).
11. Regeling van de vochtbalans :
Een homologie is gesuggereerd tussen het Golgi-complex en de contractiele vacuole van lagere Metazoa en Protozoa. De contractiele vacuole verdrijft overtollig water uit de cel. In bepaalde Protozoa houdt het Golgi-complex zich ook bezig met de regulatie van de vochtbalans.
Oorsprong van Golgi Complex:
Er zijn drie verschillende bronnen voorgesteld waaruit een nieuw Golgi-complex kan ontstaan:
1. Van endoplasmatisch reticulum :
Essner en Novik®ff (1962) en Beams en Kessel (1968) hebben voorgesteld dat de Golgi-cisternae uit de ER voortkomen. Het ruwe endoplasmatisch reticulum verliest na het synthetiseren van specifieke eiwitten ribosomen en verandert in een glad ER. Kleine vergankelijke vesicles knijpen af van een gladde ER. Deze migreren naar dictyosome.
Bij het bereiken van het vormende vlak van dictyosome smelten deze samen om nieuwe cisternae te vormen en zo bij te dragen aan de groei ervan. Door de fusie van deze blaasjes worden continu nieuwe cisternae gevormd op het vormende vlak en op het volwassen wordende vlak breken de oude cisternae af in secretoire blaasjes. Golgi vertoont dus een fenomeen van membraneuze stroming. '
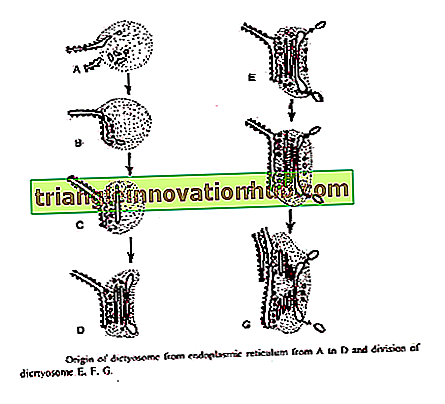

Bouch (1965) beschreef de oorsprong van Golgi van buitenmembraan van nucleaire envelop in bruine algen. Blaasjes worden afgeknepen van het buitenste kernmembraan dat samensmelt om cisternae te vormen op het vormvlak van dictyosome.
Aanwezigheid van uitsluitingszones met betrekking tot glad ER of kernmembraan, het voorkomen van zones van uitsluiting in slapende zaden van hogere planten en de vorming van dictyosome uit deze zones in ontkiemende zaden leveren bewijs voor de bovenstaande twee theorieën over de oorsprong van dictyosome.
3. Door de verdeling van reeds bestaande dictyosome :
Er is waargenomen dat tijdens celdeling in zowel planten als dieren het aantal dictyosomen toeneemt en het aantal dictyosomen in elke dochtercel net na deling bijna gelijk is aan het aantal in de oudercel voorafgaand aan deling, uit blikken en andere directe waarnemingen op de delende cellen werd verondersteld dat dictyosomen zich ook delen tijdens celdeling.